ジャパンナレッジ
アンモナイト
あんもないと
ammonites 英語
軟体動物門頭足綱に属する絶滅動物の一群の総称で、アンモン貝、菊石、菊面石ともよばれる。学術的にはアンモノイド類Ammonoideaが正しい名称である。その名は、古代エジプトの太陽神で、雄羊の頭をもつアモンΑμμο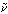 ς/Ammon(ギリシア語)に由来するが、アンモナイトの螺旋(らせん)状に巻いた殻が羊の角(つの)を連想させたからであろう。現生頭足類は1対のえらをもち、内殻ないし無殻性のイカ、タコの類(鞘形(しょうけい)亜綱Coleoidea)と、外殻性で2対のえらのオウムガイ亜綱Nautiloideaに大分類される。
ς/Ammon(ギリシア語)に由来するが、アンモナイトの螺旋(らせん)状に巻いた殻が羊の角(つの)を連想させたからであろう。現生頭足類は1対のえらをもち、内殻ないし無殻性のイカ、タコの類(鞘形(しょうけい)亜綱Coleoidea)と、外殻性で2対のえらのオウムガイ亜綱Nautiloideaに大分類される。
アンモノイド類はオウムガイ類(古生代カンブリア紀後期から現世)とよく似た多数の隔壁で仕切られた多室性の外殻を有する。しかし、発生学・比較解剖学的特徴や進化記録からはオウムガイ類と明らかに異なり、むしろ鞘形類と近縁である。かつてはアンモノイド亜綱Ammonoideaとして独立されていたが、現在ではアンモノイド目として鞘形目とともに新頭足亜綱Neocephalopodaに含める考えが有力である。アンモノイド類は、前期デボン紀(約4.2億年前)に直錐(ちょくすい)状の殻をもつバクトリテス類から分化したと考えられている。それ以降、デボン紀末期、ペルム紀末期、三畳紀末期の3回の大量絶滅事変を被りながら、世界中の海洋に大繁栄を遂げたが、白亜紀末期(約6550万年前)に陸上の恐竜類などと一緒に完全に絶滅した。その絶滅の要因については、巨大な小惑星の地球への衝突によって生じた寒冷化などの環境変動が有力視されている。
個体の構造
軟体部の大部分は不明であるが、顎器(がっき)や歯舌などの摂食器官、消化管の存在を示す化石の証拠はある。歯舌は現生鞘形類と同様、横1列が7本の小歯からなり、9本の小歯からなるオウムガイ類の歯舌と区別される。殻体は、多数の隔壁に仕切られた部屋からなる気房部と、それに続く軟体部を収容したと考えられる住房(体房)部からなる。殻を構成する螺管(らかん)は、球状ないし楕円(だえん)球状をした中空の初期室に始まり、一般には左右相称で同一平面内に巻いて成長する。初期室の後方からキチン質の管(体管)が気房部の隔壁を貫いて延びて住房部後方に開口するが、その中には動脈・静脈・結合組織・上皮(じょうひ)組織からなる軟体部が入っていたことが、アメリカ産の保存のよい化石の証拠からわかっている。連室細管の先端は初期室の手前で風船状に膨らみながら収束し、そこからへら状または管状の原体管が延びて初期室の内面に付着する。初期室から約1巻目の螺管には孵化(ふか)の際にできたと考えられるくびれがあり、そこを境に殻の構造が変化する。初期室からくびれまでの殻をアンモニテラammonitellaとよび、卵の中で形成された胚殻に相当する。アンモナイトは軟部を前方に移動させ、体管やあられ石CaCO3からなる隔壁や外殻をつくって成長した。螺管表面にはしばしば肋(ろく)、いぼ、とげ、くびれ、竜骨などの装飾を伴う。気房部の隔壁は中心から周辺に向かってオウムガイ類のものよりはるかに複雑に褶曲(しゅうきょく)し、外殻との交線の縫合(ほうごう)線suture lineは自己相似的なフラクタル曲線を描く。縫合線は個体の成長とともに複雑化するが、種類によって一定な形を示すので、重要な分類形質となる。
アンモノイド類はすべて海生、肉食性ないし腐肉食性で、その運動は、外套膜(がいとうまく)の一部である漏斗(ろうと)funnelからの海水の噴射によった。殻の内部構造の類似から、現生オウムガイ類と同様、気房部には低密度のガスと少量の水が入っていて、それにより生体の浮力や安定性を維持しながら遊泳生活をしていたと考えられる。しかし、中生代白亜紀に繁栄を遂げたニッポニテスNipponitesやハミテスHamitesなどの異常巻の類では底生に近い生活様式をもっていた可能性がある。
種類
アンモノイド類は、バクトリテス、エイゴニアタイト、アナルセステス、ゲフロセラス、クリメニア、ゴニオクリメニア、トルノセラス、ゴニアタイト、プロレカニテス(以上古生代)、セラタイト(古生代ペルム紀~中生代トリアス紀)、フィロセラス(中生代トリアス紀~白亜紀)、リトセラス、アンモナイト、アンキロセラス(以上中生代ジュラ紀~白亜紀)の14亜目に分類される。同一成長段階で比べると、縫合線の刻みは時代とともに複雑になる。
化石
アンモナイトの化石は世界各地の古生代、中生代の海成層に多産し、形態の時代的変化が著しいので、地層の時代決定や対比に有効な標準化石の代表例である。また、生物進化の研究にも広く利用されている。日本では北上(きたかみ)地方の後期古生代から白亜紀までの地層、北海道や四国の白亜紀層、山口県や福井県のジュラ紀層などから産する。とくに北海道の白亜紀層中の化石は世界的にみて保存がよく、多くの研究例がある。
©SHOGAKUKAN Inc.
