ジャパンナレッジ
遺伝子
いでんし
遺伝形質を規定する因子。遺伝子は、細胞内に存在する核酸分子、デオキシリボ核酸(DNA)の一部であり、自己複製により子孫に伝えられる。また、転写と翻訳によりタンパク質の構造を決定し、その働きにより遺伝形質を発現する。
遺伝子概念の発展
チェコのブルノの町で修道院の司祭をしていたメンデルは、1865年にエンドウの交雑実験の結果をまとめて「植物雑種についての実験」という講演を行い、翌年にこの講演の内容が論文として学会紀要に掲載された。この論文では、エンドウのそれぞれの形質に対応する細胞内の物質的なエレメント(要素)の存在が記載されていた。そして1900年に、複数の研究者がそれぞれ独立した研究によりメンデルの研究を検証することで普遍的な遺伝的法則をみいだし、遺伝学誕生の基礎となった。なお、メンデルが遺伝の仕組みを明らかにしたが長らく注目を浴びることなく、のちに彼の法則が再発見されたというのが通説である。しかし、実際にメンデルはこの論文で遺伝の仕組みについては考察しておらず、論文中のエレメント(要素)は、現在明らかになっている遺伝子という分子の存在(微小粒子の存在)を想定したものではない、という説もある。ただし、メンデルの遺伝学への貢献が否定されるものではない。
そして、遺伝を担う物質が細胞内の微小粒子であるという考えは、C・R・ダーウィンをはじめとする複数の研究者により提唱された。デンマークの遺伝学者のヨハンセンは、微小粒子の正体は不明であるとしながらも、その粒子を「ゲン(Gen)」とよぶことを提唱した。これを語源として、のちに英語では 「Gene」、日本語では「遺伝子」とよばれるようになった。
また、アメリカの生物学者であるサットンWalter Stanborough Sutton(1877―1916)が、バッタの生殖細胞を用いて染色体を観察し、遺伝の様式を染色体の性質や挙動によって説明する「染色体説」を1902年に提唱した。こうして、遺伝子と染色体の関係が明らかにされてくるなか、アメリカの遺伝学者T・H・モーガンは主としてキイロショウジョウバエを用いた研究を行い、遺伝子は染色体上に線状に配列する粒子(物質)であることを明らかにした。
1940年代には、遺伝生化学や分子遺伝学研究が発展し、遺伝子は染色体をつくる核酸の一種であり、酵素分子の働きを支配して遺伝形質を決定することが明らかにされた。1960年代には、遺伝子のもつ遺伝暗号がすべて解読され、遺伝情報の発現機構が解明された。さらに1970年代には、遺伝子の人工合成が可能になり、また細胞から取り出した遺伝子を異種の細胞に入れて増殖させ利用する遺伝子工学技術も発展してきた。
遺伝子の本体
遺伝子の本体は核酸の一種、デオキシリボ核酸(DNA)である。例外的にある種のウイルスではリボ核酸(RNA)が遺伝子として働く。
遺伝子がDNAであることが証明された科学的歴史は、1928年、イギリスの細菌学者グリフィスFrederick Griffith(1879―1941)による肺炎球菌を用いた実験に始まる。グリフィスは、肺炎球菌の細胞に含まれるなんらかの物質が、子孫に伝達される遺伝的な変化をもたらしていることを明らかにし、この現象を形質転換とよんだ。しかし、この時点ではその物質は何であるかはわかっておらず、当時、遺伝情報を運ぶのはタンパク質であると広く信じられていた。
そして1944年、アメリカの分子生物学者・細菌学者エーブリー(アベリー)、マクラウドColin Munro MacLeod(1909―1972)、マッカーティーMaclyn McCarty(1911―2005)の3人の実験により、タンパク質を分解した肺炎球菌は形質転換を起こすが、DNAを分解した肺炎球菌は形質転換を起こさないという結果が得られ、DNAが形質転換物質であると結論づけられた。
その後、1952年にアメリカの微生物学者・分子生物学者ハーシェイ(ハーシー)とチェイスがバクテリオファージT2の生活史の研究を行い、T2ファージの増殖に必要な遺伝情報をもつのはDNAであることを証明し、遺伝子がDNAの本体であることが確認された。
遺伝子DNAの構造と複製
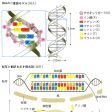
DNAは多数のデオキシリボヌクレオチドが結合してできた高分子物質である。デオキシリボヌクレオチドはリン酸、五炭糖のデオキシリボース、プリンまたはピリミジン塩基が結合したものである。DNAをつくる塩基は、アデニン(A)、グアニン(G)、シトシン(C)、チミン(T)の4種である。1950年、アメリカの生化学者シャルガフはDNA中のアデニンとチミンの数が等しく、シトシンとグアニンの数が等しいことを発見した。これらの結果は、アメリカの分子生物学者J・D・ワトソンとイギリスの物理学者・分子生物学者クリックに伝えられ、1953年に彼らがDNAの構造を発見する成果へとつながった。ワトソンとクリックが提唱したモデルは、DNA分子はデオキシリボヌクレオチドが結合してできた2本の長い鎖が螺旋(らせん)状に巻いた構造からなるというものである(図A)。2本の鎖はAとT、GとCが塩基対をつくるように結び付き、二重螺旋構造をとっている。二重螺旋の直径は2.0ナノメートル、螺旋の1回転の距離は3.4ナノメートルで、その間に塩基対が10個並んでいる。このようなDNA鎖をつくる塩基の配列順序は遺伝暗号として働き、遺伝情報を決定するものである。DNAの量は種によって異なるが、遺伝学研究によく用いられる大腸菌の全DNA塩基配列は464万塩基対であり、約4000の遺伝子をもつと推定されている。
遺伝子は細胞分裂の際に正しく同じものに複製され、子孫細胞に伝えられる。DNA分子の複製の際には、二重螺旋構造は部分的に1本鎖となり、それぞれの鎖を鋳型として、DNA合成酵素の働きにより対になる新しい鎖が合成される。このようにDNA分子の複製は自己複製であり、また二重鎖の1本から新しい分子がつくられるので半保存的複製とよばれる。
遺伝子と染色体
高等動植物など真核生物の細胞では、DNAはヒストンタンパク質に巻きつけられたヌクレオソームnucleosomeという構造をとっている。多数のヌクレオソームが連なった構造をクロマチンchromatinとよび、染色体はこのクロマチンが折り畳まれて高次構造をとったものである。狭義には、細胞の分裂時にクロマチンが凝縮して棒状の構造をとったものを染色体とよぶが、広義には、その形や細胞の分裂周期にかかわらず、クロマチンの構造体を染色体とよぶ。細菌類など原核生物の細胞では、細胞分裂の際に真核細胞でみられるような核や染色体をつくらず、DNA分子はクロマチン構造をとることなく細胞質中に分布している。
遺伝子は染色体上に線状に配列している。二つの遺伝子が異なる染色体上にあるときには、次世代への継承はメンデルの独立の法則に従って分離するが、同じ染色体上にあるときにはこの法則に従わず、行動をともにし、連鎖の現象を示す。1本の染色体上の遺伝子は一つの連鎖群を形成するが、交雑の結果、連鎖している遺伝子の組合せが親と異なる組合せに変わることがあり、この現象は遺伝的組換えとよばれる。組換えは減数第1分裂の過程で対合した相同染色体の間で交差と乗換えが起こり、新しい遺伝子の組合せとなる組換え型が生じる現象である。組換え型の出現頻度をパーセント(%)で示した値を組換え価とよぶ。組換え価を遺伝子間の距離とし、これを線上に目盛ると、遺伝子が染色体上にどのように並んでいるかを示す図ができる。この図は染色体地図、遺伝地図、あるいは連鎖地図とよばれる。
組換えはDNA分子間の交差切断と、相同な相手分子への乗換えによって起こると考えられている。組換え過程では、DNA鎖の切断酵素、修復酵素、連結酵素などが働いている。DNA鎖の特定塩基配列部位を切る酵素は制限酵素とよばれる。同じ制限酵素で切った2種のDNA鎖は切り口の構造が相補的であり、それらを結合してできた分子は組換えDNAとよばれる。宿主(しゅくしゅ)細胞内で自己増殖する核外遺伝子プラスミドのDNAと目的とする特定の遺伝子DNAとの間で組換えDNAをつくり、それを宿主細胞に導入することで遺伝子のコピーを増やすことができる。これは遺伝子のクローン化とよばれる現象で、遺伝子工学の主要な手段となっている。
遺伝子の働き
遺伝子は遺伝形質を決定する。アメリカの遺伝学者のビードルとテータムは1941年に特定の栄養分を必要とするアカパンカビの突然変異体の研究を行い、「一遺伝子一酵素説」を提唱した。この仮説は、遺伝子は一つの酵素の構造や働きを支配し、遺伝形質を発現するとするものである。その後、1958年にクリックが分子生物学の「セントラルドグマ」(中心教義)として主張しているように、遺伝子DNAの遺伝情報はまず伝令RNAに転写されて、細胞質に移動する(図B)。伝令RNAはリボゾームとよばれる小粒に付着し、運搬RNAなどの働きで翻訳され、翻訳されたものから遺伝暗号に従ってアミノ酸が結合され、最終的に結合されたものからタンパク質のポリペプチド鎖が合成される(図B)。タンパク質は酵素として細胞内の代謝反応を触媒し、また細胞構造をつくり遺伝形質を発現させる。
遺伝子のもつ遺伝暗号は、1961年から約5年間をかけて解読された。解読された遺伝暗号は、遺伝子DNAの三つのヌクレオチドがコドンcodonとよばれる暗号の単位となって一つのアミノ酸を指定するというもので(図B)、トリプレット暗号ともいわれる。コドンは64種あり、そのうち61種はタンパク質をつくる20種のアミノ酸のどれかを指定するが、3種のコドンはどのアミノ酸も指定せず、遺伝暗号の読みの終止暗号として働く。また、必須アミノ酸メチオニンの暗号(AUG)は遺伝暗号の読みの開始暗号となる。ウイルスからヒトに至るまで、どの生物も同じ遺伝暗号を用いている。
遺伝子が働くか働かないかは、環境条件や遺伝子がもつ調節機能により制御されている。フランスの分子遺伝学者F・ジャコブと分子生物学者J・L・モノーは1961年に大腸菌の乳糖代謝の調節機構の研究から、酵素合成が調節遺伝子によりオペロンとよばれる遺伝子群を単位として調節されるという「オペロン説」を提出した。オペロン説によると、大腸菌の培養中に乳糖がないときには、調節遺伝子からつくられる調節物質(リプレッサーとよばれるタンパク質)がオペロンの一端(オペレーターとよばれる)に結合し酵素合成を止めているが、乳糖が加えられるとリプレッサーは不活性化され、オペロンから酵素が合成される。真核細胞は多数の遺伝子をもつが、組織に特有の遺伝子のみが働き、ほかは働きを停止している。このような真核細胞における遺伝子作用調節機構は、染色体構造と密接な関係をもつものと考えられている。
突然変異
DNA複製の際のミス、動く遺伝子(転移因子)トランスポゾンの転移、物理的要因によるDNAの損傷などにより、DNA配列に起きる変化を突然変異とよぶ。物理的要因としては、放射線の照射や、アルキル化剤のようなDNAに作用する化学物質などが知られている。突然変異はDNA塩基対の置換、欠失、転座、逆位、挿入などにより、遺伝子配列上に変異が生じ、指定するアミノ酸配列が変わることによって起こる。突然変異で遺伝子機能の性質が変化することで、生存に有利に働いたり、逆に遺伝子機能が失われたりする。
遺伝子記号と種類
遺伝子の名称は遺伝子記号で表される。遺伝子記号は、その遺伝子の決定する形質の特徴を示す英語やラテン語などの省略形と、番号や記号からつくられる。たとえば、微生物でアミノ酸の一種であるトリプトファンがないと生育できない形質を示す遺伝子の記号はtrpである。トリプトファン合成系にはいくつもの酵素反応があり、それぞれの反応を支配する遺伝子には番号やアルファベットをつけtrp1、trp2あるいはtrpA、trpBのようによぶ。DNAやRNAをさす場合はイタリック(斜体)、タンパク質をさすときはローマン(正体)文字とする。また、遺伝子とアレル(対立遺伝子)の記載は区別する必要があるが、混同されている場合も多い。遺伝子の記載方法は生物種によって異なるが、アレルの記載は、潜性アレルを小文字、顕性アレルを大文字で表す。
遺伝子の固有名とは別に、遺伝子が細胞のどの部分にあるか、どのような形質を支配するか、どのような働きをするかなどにより、種類分けがなされている。核にある遺伝子は核遺伝子あるいは染色体遺伝子とよばれる。細胞質にあって細胞質遺伝の原因となるものは細胞質遺伝子とよばれ、とくに細胞質中の細胞小器官であるミトコンドリアに存在するものはミトコンドリア遺伝子とよばれる。一遺伝子一酵素説に従いタンパク質の構造を決定しているのは構造遺伝子であり、構造遺伝子の働きを誘導したり抑制したりしているのが調節遺伝子である。また、生物個体の発生過程で致死作用を示す遺伝子は致死遺伝子とよばれる。自然界に普通にみられる正常な遺伝子は野生型遺伝子で、変化すると突然変異型遺伝子になる。また、一つの形質を発現するのに2種類以上の異なる遺伝子座にある遺伝子が働いていることがあり、これらの遺伝子は同義遺伝子とよばれる。これらのほかにも、各種の遺伝現象を表すために、多数の名称が記載されている。
©SHOGAKUKAN Inc.