
《同》濾過性病原体(filterable microorganism).DNAかRNAのどちらかをゲノムとしてもち,細胞内だけで増殖する感染性の微小構造体.ラテン語で毒(virus)を意味し,後に転じて病原体を意味するようになった.D.I.Ivanovski(1892)はタバコモザイク病が細菌濾過器を通した濾液で感染することを観察,F.LoefflerとP.Frosch(1898)は口蹄疫が同じく細菌濾過器を通過した濾液で感染することを見出し,これをultramicroscopic filterable virusesと記載した.M.W.Beijerinck(1898)は,この濾過性病原体は微小な細菌ではなく,未知の溶液状の物体であると主張し,これをContagium vivum fluidumと呼んだ.これらの研究がウイルスの発見として位置づけられている.ウイルス粒子を確認する技術の発達,例えば電子顕微鏡の進歩やウイルスの培養技法の確立以前には,ウイルスに因子(factor)という無生物的な呼称を用いたこともある(例:乳因子).A.M.Lwoffら(1957)は,細胞内だけで増殖し,潜在的に病原性をもつ感染性の実体のうち,次のような属性をもつものをウイルスと定義した.(1)核酸としてDNAかRNAのどちらか一方をもつ,(2)遺伝物質(核酸)だけから複製される,(3)二分裂で増殖しない,(4)エネルギー産生系を欠く,(5)宿主のリボソームを蛋白質合成に利用する.これらのことから一般に生物とは別の扱いをされる.その起原については,小型のトランスポゾンが独立して細胞外に出られる機能をそなえるに至ったものとする説など,諸説がある.ゲノムがRNAのものをRNAウイルス(RNA virus),DNAのものをDNAウイルス(DNA virus)と呼ぶ.RNAゲノム,DNAゲノムとも一本鎖,二本鎖のものがあり,ゲノムの性質に応じて複製様式が異なる.植物ウイルスはほとんどがRNAウイルスである.ウイルスを研究する学問分野をウイルス学(virology)という.(⇒ウイロイド,⇒ウイルスの分類,付録:ウイルス分類表)
ビールス,バイラスなどとも発音される。核酸(DNAもしくはRNA)とタンパク質からなる,細菌よりも小さな一群の病原体。遺伝情報を担う核酸がタンパク質の外被におおわれた構造をもち,それぞれのウイルスに特有の宿主となる細菌や生物の細胞に寄生して,宿主のタンパク質合成能やエネルギーを利用して,自己増殖を行う。ウイルスは,その大きさが数十~数百nmときわめて小さく単純であることと,単独では生物としての要件である自己増殖能をもたず,寄生してはじめて自己増殖を行うことから,しばしば〈生物と無生物の間にあるもの〉と表現されることがある。また,ワクチンによる免疫療法を除くと,細菌などに対する抗生物質のように,ウイルス感染症に対する直接的な特効薬がいまだに発見されていないのも,ウイルスの独特な寄生性と自己増殖性によるものである。
19世紀に入って,L.パスツールやR.コッホたちの研究により,伝染病の多くが病原性細菌によってひき起こされていることが明らかにされた。しかし,伝染病のうちには,これらの病原性細菌が見いだされないものがあった。1892年にイワノフスキーD.I.Ivanovskiiがタバコモザイク病で,98-99年にレフラーF.LoefflerとフロッシュP.Froschがウシの口蹄疫で,それぞれ,細菌を通さない細菌ろ過器を通過させたろ液それ自体が,明確に病原性をもつことを示した。引き続いて,黄熱,狂犬病,牛痘などの伝染病の病原が,同じように細菌ろ過器を通過することが確認され,20世紀の初頭には〈ろ過性病原体filterable virus〉という概念が確立された。ウイルスという言葉はもともとはラテン語の〈毒素〉という意味で,一般に伝染病の病原因子を総称していたが,1930年代以降は,ろ過性病原体を意味するようになった。
コッホによって確立された細菌学の特徴的な手法は,培地による細菌の培養であった。初期のウイルス研究においても,さまざまな培地でウイルスを培養する試みがなされ,30年代が最も盛んであった。しかし,このような試みは結局失敗に終わった。ウイルスを分離培養するためには,適切な生きた細胞が必要とされている。現在,ウイルスの分離培養に用いられている手段は以下のようである。(1)実験動物による培養,(2)発育鶏卵(孵化(ふか)の途中の受精卵)による培養,(3)培養細胞による培養。しかし現在においても,適切な培養手段が見いだせないために研究の進んでいないウイルスも存在している。
30年代までのウイルス学は,培地による培養という細菌学の手法を適用できないという困難をかかえたまま,病原性細菌学の一部として取り扱われていた。しかしこの時期以降,ウイルス学は現在の分子生物学の発展の基盤を形づくるようになってくる。35年スタンリーW.M.Stanleyはタバコモザイクウイルスの結晶化に成功したが,このことは,結晶化したウイルスが生物か無生物かという論議をひき起こした。この問いに答えることは今日でも難しいが,スタンリーの研究は,遺伝情報の担い手である遺伝子の化学的本体の追求へと引き継がれていった。遺伝子の本体がDNAであることがはっきり証明されたのは,52年のハーシェーA.D.HersheyとチェースM.Chaseによる大腸菌に感染するT2ファージを用いた実験によってであった。そして53年のJ.D.ワトソンとF.H.C.クリックによるDNAの二重らせん構造の解明を契機として,分子生物学の飛躍的な発展がもたらされる。50年代に取り扱われたウイルスはバクテリオファージが中心であったが,60年代以降,これらの分子生物学の知見と,培養細胞によるウイルスの培養方法の確立とともに,動物ウイルスにも研究の目が注がれてきている。今日,ウイルス学が取り扱う範囲は,ウイルス自体についての形態形式,遺伝子構造と遺伝子の機能発現などだけにとどまらず,宿主細胞の側での遺伝子の構造と機能,発癌やウイルスの病原性を定めている遺伝子とその機能,ウイルスに対する防御機構などの研究にも及んでいる。
自己増殖していくための遺伝情報は,ウイルスにおいてもまた核酸によって担われているが,高等生物の遺伝情報はDNAに限られるのに対して,ウイルスの場合にはRNAのときもある。ウイルス自体の大きさが微小なため,遺伝子としてのウイルスの核酸のもつ情報量は制限されており,核酸の情報に基づいて生み出されるタンパク質の種類も限られている。このような制限のもとでつくり出されてくるウイルス粒子は,基本的には,単位的な要素(サブユニット)の規則的な繰返しによって構成されている。サブユニットは多くの場合,同一のタンパク質の集合したものであるが,数種類のタンパク質からなる複雑な集合体である場合もある。
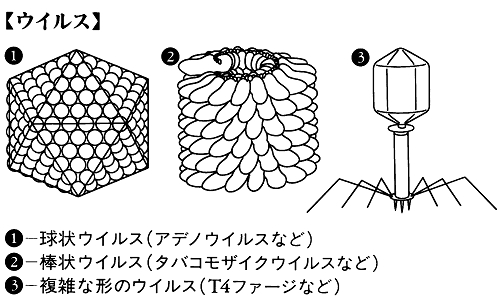
ウイルスの基本的な形態は,遺伝子である核酸をタンパク殻(カプシドcapsid)でおおっている形である。ウイルスカプシドを構成しているタンパク質サブユニットの配列方法は,正二十面体からせん構造になっている。限られた種類のサブユニットからカプシドをつくると,正二十面体からせん構造になるのである。したがって,電子顕微鏡でウイルスを拡大して見た場合,ウイルス粒子の形状は正二十面体を反映した球状,らせん構造を反映した棒状,およびこれらの形の複合した複雑な形の三つに大別されることが観察されている。複雑な形のウイルス(T4ファージなど)では,ウイルス粒子を構成しているタンパク質の種類数も多くなっており,サブユニットの種類数も多くなっている。
ある種のウイルスでは,カプシドの外側にエンベロープenvelopeと呼ばれる膜様構造物をもっている。エンベロープは,タンパク質,脂肪,炭水化物などを含んでいる。
ウイルスは,それが感染する宿主の種類によって,動物ウイルス,植物ウイルス,昆虫ウイルス,細菌ウイルス(バクテリオファージ)の四つに大別される。
1960年代以降の電子顕微鏡やX線回折によるウイルス形態学の進歩および分子生物学の発展によって,ウイルスの一元的な分類体系がつくられつつある。分類の基準となるものは,ウイルスのもつ核酸がDNAかRNAか,カプシドの形状,エンベロープの有無,ウイルス粒子が形成される宿主細胞中での位置などである。
人体に侵入したウイルスは,さまざまな病気を引き起こす。ウイルス性の疾患には,胃腸炎,肝炎,出血熱,腫瘍,髄膜炎,肺炎,皮膚疾患などが知られている。
ウイルス性肝炎は,肝炎ウイルスの感染による。肝炎ウイルスはいろいろなウイルスの総称で,A~E型までの,少なくとも5種類の肝炎ウイルスが確認されている。A,C,D,E型は核酸がRNAのRNAウイルスで,B型は核酸がDNAのDNAウイルスである。ウイルス性出血熱には,黄熱,デング熱,ラッサ熱,エボラ出血熱,マールブルグ病などがあり,これらの病原体はRNAウイルスである。ラッサ熱,エボラ出血熱,マールブルグ病の三つは,予防法や治療法が確立されていず,致命率が高く,伝染力も強いことで知られている。ウイルス性腫瘍には,Epstein-Barrウイルス(EBV)によるバーキットリンパ腫と鼻咽頭癌,ヒト白血病ウイルス(HTLV)による成人T細胞白血病が知られている。EBVはDNAウイルスであり,HTLVはRNAウイルスである。
エイズ(後天性免疫不全症候群)はエイズウイルス(ヒト免疫不全ウイルス,HIV)の感染による。細胞性免疫不全により,日和見感染症,カポジ肉腫,リンパ腫などが引き起こされる。エイズウイルスはRNAウイルスで,HTLVとともにレトロウイルスと呼ばれるRNAウイルスの仲間である。
→癌
ウイルスが感染した細胞では,細胞内でウイルス核酸の情報に基づいて,タンパク質と核酸の合成が行われ,その結果,新たに数多くのウイルスが出現することになる。この過程が繰り返されていくと,生体の側には,そのウイルスに特有の感染症状が現れる。ウイルスに対して生体側がとる防御方法は,インターフェロン系と免疫系の働きによる2種類がある。インターフェロン系の働きは,ウイルス感染の初期にインターフェロンを産生し,生体の細胞を抗ウイルス状態にさせ,ウイルスの細胞内での増殖を抑えることにある。免疫系の働きには,抗体がつくられる液性免疫と,免疫細胞が働く細胞性免疫の2種類がある。液性の免疫系では,ウイルス粒子が非自己の物質として生体の免疫系に認識され,そのウイルスに特異的な抗体がつくり出される。抗体はウイルスと結合して,ウイルスの感染力を失わせる。細胞性の免疫系では,ウイルスが感染したことによって本来の機能を失った細胞を免疫細胞が攻撃することによって,それらの細胞を生体からとり除く。
→バクテリオファージ
植物に感染するウイルスを植物ウイルスと呼び,おもに高等植物に病気を起こすものを指すが,菌類(キノコを含む)や藻類に感染するものもある。植物ウイルスは形,大きさ,宿主範囲と病徴,伝染方法,核酸の性状などに基づき約600種が同定されているが,それらは約30のグループに分類される。ウイルスによって起こる病徴はウイルスの種類によりさまざまであるが,最もふつうにみられるのは葉の緑色に濃淡のまだらが生じるモザイクで,葉脈に沿うこともあり,単子葉植物では縞になることが少なくない。ほかに,黄化,壊死(えし),輪紋,葉巻,萎縮,奇形などがある。これらの病徴は葉,茎,花,果実の各器官に現れるが,根は通常発達が悪くなる。果樹や樹木では,はっきりした病徴が認められないことも多い。植物体内では動物と異なり抗体をつくる機能がないのでウイルスに対する免疫ができず,一般にウイルスは植物が枯れるまで残存する。ウイルスが増殖する細胞・組織はウイルスによって異なるが,どのウイルスも生長点付近のごく一部を除いて植物の全身に分布するため,株分け,挿芽,球根など栄養繁殖する植物では,母株のウイルスはそのまま苗に伝達される。接木をすると組織の癒合によりウイルスは必ず伝染する。一方,種子伝染性のウイルスは,マメ類のウイルス病やセンチュウ媒介のウイルス病など一部にみられる程度で,伝染率も平均してそう高くない。
野外における最も重要な伝染方法は昆虫による媒介(虫媒伝染ともいう)である。アブラムシ,ヨコバイ,ウンカ類は口針で植物汁液を吸収して害を与えるだけでなく,吸汁の際にウイルスを伝染する機会が多い。畑作物,野菜,花のウイルス病の多くはアブラムシで媒介される。キュウリモザイクウイルス,ジャガイモYウイルスなどはアブラムシの短時間の吸汁で伝染するが,虫体内でウイルスが保持される時間は短い。イネのウイルス病にはヨコバイまたはウンカで媒介されるものが多く,イネ萎縮病ではヨコバイが病植物を吸汁するとウイルスが昆虫細胞内で増殖し,その虫は終生伝染を続ける。さらに,雌虫が保毒すると卵を通して子虫へ高率にウイルスが伝染するが,これを経卵伝染という。その他,アザミウマ,コナジラミ,コナカイガラムシ,ハムシなどの昆虫やダニによって媒介されるウイルスがある。センチュウや遊走子をもつ下等菌類によって媒介されるウイルスは,それらが生息する土壌を通して伝染する。実験的には多数のウイルスが汁液で伝染するが,農作業の際などに接触して伝染するウイルスはタバコモザイクウイルスなど一部にすぎない。ウイルス病の防除には伝染を断つことが最も肝要であるが,組織培養による無病苗の生産も実効を上げており,また一部では病原性の弱い変異ウイルスの干渉効果を利用した方法も行われる。



 [リ]
[リ]


©2026 NetAdvance Inc. All rights reserved.